
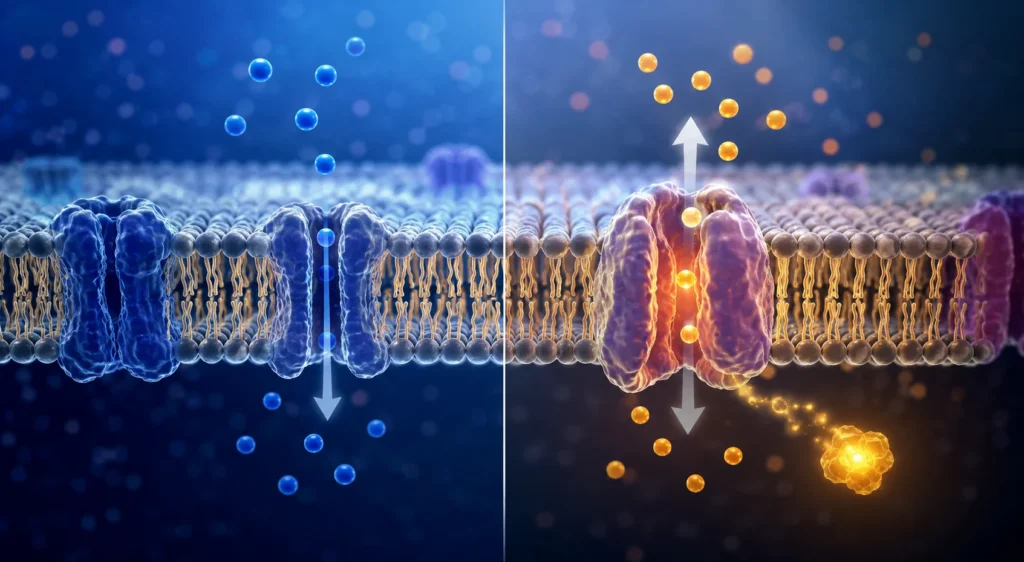
La supervivencia celular depende de la capacidad de la membrana plasmática para regular de manera precisa el intercambio de sustancias entre el medio intracelular y el medio extracelular. Esta regulación permite mantener la homeostasis, conservar gradientes electroquímicos, controlar el volumen celular, obtener nutrientes esenciales, eliminar productos de desecho y coordinar innumerables procesos fisiológicos. A pesar de la enorme diversidad de moléculas que atraviesan las membranas biológicas, los mecanismos fundamentales mediante los cuales ocurre dicho transporte pueden agruparse en dos categorías principales: la difusión y el transporte activo. Ambos procesos comparten el objetivo general de movilizar sustancias a través de una barrera biológica, pero difieren profundamente en la fuente energética que utilizan, en la dirección del movimiento molecular respecto a los gradientes energéticos y en las consecuencias fisiológicas derivadas de su funcionamiento.
La membrana plasmática está constituida principalmente por una bicapa de fosfolípidos en la cual se encuentran insertadas numerosas proteínas integrales y periféricas. Esta organización genera una barrera semipermeable que restringe el libre movimiento de la mayoría de las moléculas hidrosolubles, permitiendo al mismo tiempo una regulación altamente selectiva del intercambio de sustancias. Las propiedades físicas y químicas de esta estructura determinan la necesidad de mecanismos especializados de transporte que permitan el desplazamiento controlado de moléculas entre ambos lados de la membrana. Esta concepción estructural de la membrana y su papel como barrera selectiva han sido demostrados mediante estudios biofísicos, estructurales y moleculares desarrollados durante varias décadas de investigación científica.
La difusión representa el mecanismo más elemental de transporte transmembranal. Este proceso se basa en el movimiento aleatorio continuo de las moléculas como consecuencia de su energía térmica intrínseca. Todas las moléculas presentes en un sistema biológico experimentan un movimiento constante debido a la energía cinética asociada a la temperatura. Como resultado, las partículas colisionan entre sí y cambian continuamente de dirección. Cuando existe una diferencia de concentración entre dos regiones, el movimiento aleatorio produce un flujo neto de partículas desde la región donde la concentración es mayor hacia aquella donde la concentración es menor. Aunque cada molécula individual se desplaza al azar, el comportamiento colectivo de millones de moléculas genera una transferencia neta predecible hacia el equilibrio.
La difusión constituye un proceso espontáneo porque ocurre en la dirección que disminuye la energía libre del sistema. El gradiente de concentración representa una forma de energía potencial. Cuando las moléculas se distribuyen uniformemente, el sistema alcanza un estado de equilibrio termodinámico caracterizado por una mayor entropía y una menor energía libre. Por esta razón, la difusión no requiere una fuente metabólica adicional de energía, ya que la energía necesaria para el desplazamiento molecular proviene directamente de la energía térmica existente en el sistema.
La velocidad de difusión depende de múltiples factores físicos. Entre ellos destacan la magnitud del gradiente de concentración, la temperatura, el tamaño molecular, la viscosidad del medio, la distancia de difusión y las características estructurales de la membrana. Un aumento del gradiente de concentración incrementa la fuerza impulsora del movimiento neto. De manera similar, un incremento de la temperatura aumenta la energía cinética molecular y favorece una mayor frecuencia de colisiones, acelerando el proceso difusivo. Por el contrario, las moléculas de gran tamaño suelen difundirse más lentamente debido a sus menores coeficientes de difusión.
La difusión puede clasificarse en difusión simple y difusión facilitada. La difusión simple ocurre cuando las moléculas atraviesan directamente la bicapa lipídica o pasan a través de poros acuosos sin necesidad de un mecanismo específico de transporte. Las sustancias liposolubles, como el oxígeno, el dióxido de carbono, los esteroides y numerosos anestésicos, atraviesan fácilmente la región hidrofóbica de la membrana mediante este mecanismo. Debido a su elevada solubilidad en lípidos, estas moléculas pueden disolverse temporalmente en la bicapa y desplazarse a través de ella impulsadas exclusivamente por sus gradientes de concentración.
El oxígeno constituye un ejemplo clásico de difusión simple. La presión parcial de oxígeno es mayor en los alvéolos pulmonares que en la sangre venosa que llega a los capilares pulmonares. Esta diferencia genera un gradiente que impulsa la difusión espontánea de oxígeno hacia la sangre. Posteriormente, otro gradiente favorece el paso del oxígeno desde los capilares sistémicos hacia los tejidos. Todo este proceso ocurre sin gasto metabólico directo y constituye una manifestación fundamental de la difusión pasiva en los organismos multicelulares.
La difusión facilitada también depende de gradientes de concentración y continúa siendo un proceso pasivo, pero requiere la participación de proteínas especializadas de membrana. Estas proteínas pueden actuar como canales o como transportadores. Los canales proporcionan una vía acuosa continua a través de la membrana, permitiendo el paso rápido y selectivo de determinadas sustancias. Los transportadores, por su parte, se unen específicamente al soluto y experimentan cambios conformacionales que facilitan su transferencia de un lado al otro de la membrana.
La difusión facilitada resulta indispensable para el transporte de muchas moléculas hidrosolubles que no pueden atravesar libremente la bicapa lipídica. La glucosa constituye uno de los ejemplos más estudiados. En numerosos tejidos, la glucosa penetra en las células mediante proteínas transportadoras pertenecientes a la familia GLUT. Estas proteínas reconocen específicamente la molécula de glucosa y permiten su desplazamiento siguiendo su gradiente de concentración. Aunque intervienen proteínas especializadas, el proceso continúa siendo pasivo porque el movimiento ocurre a favor del gradiente electroquímico de la glucosa y no requiere energía metabólica directa.
Una característica distintiva de la difusión facilitada es la saturación. Debido a que el número de proteínas transportadoras es finito, existe una velocidad máxima de transporte. Cuando todos los transportadores disponibles se encuentran ocupados, un incremento adicional de la concentración del soluto no produce un aumento proporcional en la velocidad de transporte. Este comportamiento cinético recuerda al observado en las reacciones enzimáticas y refleja la naturaleza específica de las interacciones entre transportadores y sustratos.
A diferencia de la difusión, el transporte activo permite mover sustancias en contra de sus gradientes electroquímicos. Este fenómeno posee una enorme importancia fisiológica porque posibilita la acumulación selectiva de moléculas en compartimentos celulares específicos. Sin transporte activo, las concentraciones intracelulares y extracelulares tenderían gradualmente a igualarse, lo que comprometería funciones esenciales como la excitabilidad neuronal, la contracción muscular, la secreción glandular y el transporte epitelial.
El movimiento de una sustancia en contra de su gradiente implica un aumento de la energía libre del sistema. Por esta razón, dicho proceso no puede ocurrir espontáneamente y requiere el aporte de energía procedente de una fuente externa. En la mayoría de los casos, esta energía deriva directa o indirectamente de la hidrólisis de adenosín trifosfato.
El transporte activo depende de proteínas transportadoras especializadas conocidas como bombas o transportadores activos. Estas proteínas experimentan cambios conformacionales impulsados por reacciones energéticas que permiten desplazar solutos contra gradientes de concentración o gradientes eléctricos. La especificidad molecular de estos sistemas es extraordinariamente elevada y permite a las células controlar con precisión la composición química de su medio interno.
El transporte activo puede dividirse en transporte activo primario y transporte activo secundario. En el transporte activo primario, la energía procede directamente de la hidrólisis de adenosín trifosfato. Las proteínas responsables poseen actividad adenosín trifosfatasa y utilizan la energía liberada durante la hidrólisis para impulsar cambios estructurales que permiten el movimiento del soluto.
La bomba sodio-potasio constituye el ejemplo paradigmático de transporte activo primario. Esta proteína transporta tres iones sodio desde el interior celular hacia el exterior y dos iones potasio desde el exterior hacia el interior durante cada ciclo de transporte. Como resultado, mantiene concentraciones elevadas de potasio dentro de la célula y concentraciones elevadas de sodio en el espacio extracelular. Este gradiente es esencial para la generación del potencial de membrana en reposo, la conducción nerviosa, la contracción muscular, el transporte de nutrientes y la regulación osmótica celular.
La importancia fisiológica de esta bomba es extraordinaria. Se estima que una fracción significativa del consumo energético total de muchas células se destina al funcionamiento continuo de la bomba sodio-potasio. En neuronas, por ejemplo, este sistema representa uno de los principales consumidores de energía metabólica debido a la necesidad constante de mantener gradientes iónicos compatibles con la transmisión de impulsos nerviosos.
Otros ejemplos de transporte activo primario incluyen las bombas de calcio y las bombas de protones. Las bombas de calcio mantienen concentraciones citoplasmáticas extremadamente bajas de este ion, condición indispensable para la señalización intracelular. Las bombas de protones participan en la acidificación de compartimentos intracelulares y en la secreción de ácido gástrico por las células parietales del estómago.
El transporte activo secundario utiliza una estrategia diferente. En este caso, la energía no procede directamente de la hidrólisis de adenosín trifosfato en el propio transportador. En cambio, aprovecha la energía almacenada en gradientes electroquímicos previamente establecidos por sistemas de transporte activo primario. El gradiente de sodio generado por la bomba sodio-potasio constituye la fuente energética más frecuente para este tipo de transporte.
Cuando los iones sodio tienden a ingresar nuevamente a la célula siguiendo su gradiente electroquímico, la energía liberada durante ese movimiento puede utilizarse para transportar simultáneamente otra sustancia contra su propio gradiente. Este mecanismo permite el transporte eficiente de nutrientes y otros solutos esenciales.
Un ejemplo clásico es el cotransportador sodio-glucosa presente en el epitelio intestinal y en los túbulos renales. Este sistema aprovecha la tendencia natural del sodio a entrar en la célula para impulsar la captación de glucosa incluso cuando la concentración intracelular de glucosa es superior a la concentración luminal. Gracias a este mecanismo, el organismo puede absorber eficazmente nutrientes desde el tracto digestivo y recuperar glucosa filtrada en el riñón.
La comparación entre difusión y transporte activo revela diferencias fundamentales en términos energéticos, funcionales y fisiológicos. La difusión siempre ocurre a favor de un gradiente de energía y no requiere gasto metabólico adicional. El transporte activo, por el contrario, desplaza sustancias contra gradientes electroquímicos y requiere una fuente externa de energía. La difusión tiende a eliminar diferencias de concentración y conducir al equilibrio, mientras que el transporte activo genera y mantiene dichas diferencias. La difusión aprovecha gradientes preexistentes; el transporte activo crea gradientes que posteriormente pueden ser utilizados para múltiples funciones celulares.
Ambos mecanismos no son procesos independientes, sino componentes complementarios de una estrategia fisiológica integrada. El transporte activo establece gradientes electroquímicos mediante gasto energético, mientras que la difusión permite aprovechar esos gradientes para realizar trabajo biológico útil. La interacción continua entre ambos procesos constituye uno de los principios fundamentales de la fisiología celular y explica cómo las células mantienen un entorno interno organizado, dinámico y compatible con la vida.


Fuente y lecturas recomendadas:
Alberts, B., Johnson, A., Lewis, J., Morgan, D., Raff, M., Roberts, K., & Walter, P. (2022). Molecular biology of the cell (7th ed.). Garland Science.
Berg, J. M., Tymoczko, J. L., Gatto, G. J., & Stryer, L. (2019). Biochemistry (9th ed.). W. H. Freeman.
Hille, B. (2001). Ion channels of excitable membranes (3rd ed.). Sinauer Associates.
Jardetzky, O. (1966). Simple allosteric model for membrane pumps. Nature, 211(5052), 969–970. https://doi.org/10.1038/211969a0
Karp, G. (2022). Cell and molecular biology: Concepts and experiments (9th ed.). Wiley.
Lodish, H., Berk, A., Kaiser, C. A., Krieger, M., Bretscher, A., Ploegh, H., Amon, A., & Martin, K. C. (2021). Molecular cell biology (9th ed.). W. H. Freeman.
Mitchell, P. (1961). Coupling of phosphorylation to electron and hydrogen transfer by a chemi-osmotic type of mechanism. Nature, 191(4784), 144–148. https://doi.org/10.1038/191144a0
Nelson, D. L., & Cox, M. M. (2021). Lehninger principles of biochemistry (8th ed.). W. H. Freeman.
Singer, S. J., & Nicolson, G. L. (1972). The fluid mosaic model of the structure of cell membranes. Science, 175(4023), 720–731. https://doi.org/10.1126/science.175.4023.720
Skou, J. C. (1957). The influence of some cations on an adenosine triphosphatase from peripheral nerves. Biochimica et Biophysica Acta, 23, 394–401. https://doi.org/10.1016/0006-3002(57)90343-8
Ussing, H. H. (1949). The distinction by means of tracers between active transport and diffusion. Acta Physiologica Scandinavica, 19(1), 43–56. https://doi.org/10.1111/j.1748-1716.1949.tb00602.x
Zheng, J., & Trudeau, M. C. (Eds.). (2015). Handbook of ion channels. CRC Press.
Síguenos en X: @el_homomedicus y @enarm_intensivo Síguenos en instagram: homomedicus y en Treads.net como: HomomedicusAprende administración paso a paso

ADMINISTRACION DESDE CERO
