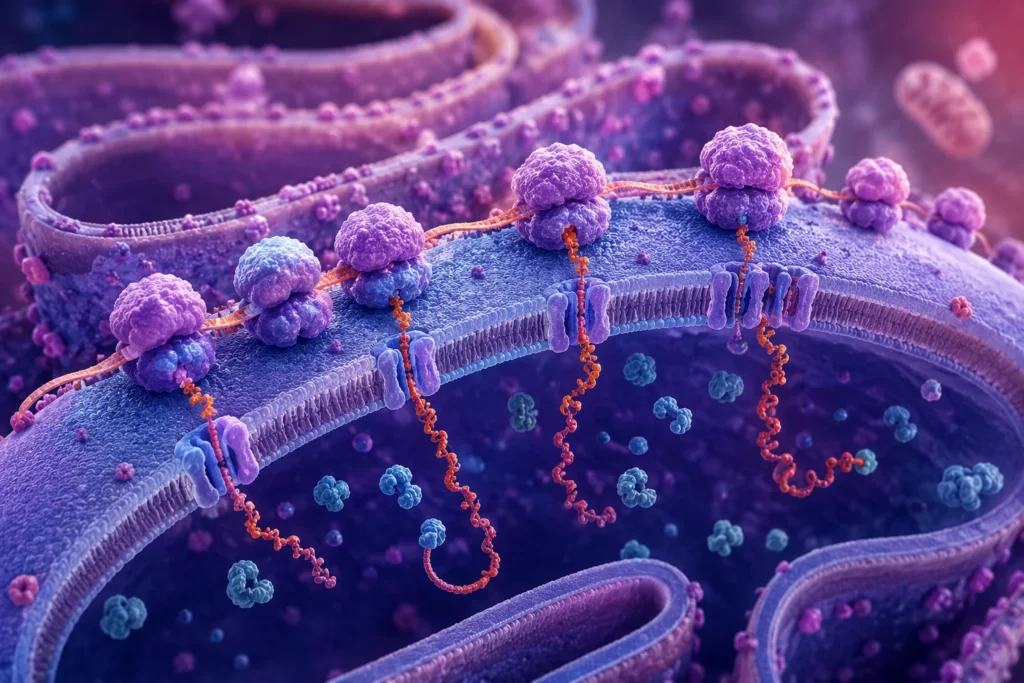
La adhesión de numerosos ribosomas al retículo endoplásmico constituye uno de los procesos fundamentales de la biología celular eucariótica, ya que permite la síntesis, el procesamiento inicial y la distribución intracelular de una gran proporción de las proteínas producidas por la célula. Esta asociación no ocurre de manera aleatoria, sino que depende de mecanismos moleculares altamente regulados que garantizan que cada proteína alcance el compartimento celular apropiado desde los primeros momentos de su síntesis.
Los ribosomas citoplasmáticos son estructuras dinámicas que pueden encontrarse libres en el citosol o asociados transitoriamente a la membrana del retículo endoplásmico. Ambos grupos poseen una estructura esencialmente idéntica; sin embargo, su localización durante la traducción depende de la información codificada en la propia proteína naciente. Las proteínas destinadas al citosol completan su síntesis sobre ribosomas libres, mientras que aquellas dirigidas al sistema de endomembranas, a la membrana plasmática, a los lisosomas o a la secreción extracelular son sintetizadas inicialmente por ribosomas libres que posteriormente son reclutados hacia la superficie del retículo endoplásmico mediante señales específicas presentes en la cadena polipeptídica en crecimiento.
El elemento determinante de este proceso es la denominada secuencia señal, una región localizada habitualmente cerca del extremo amino terminal de la proteína naciente. Esta secuencia está formada por un conjunto característico de aminoácidos hidrofóbicos capaces de ser reconocidos por un complejo ribonucleoproteico especializado denominado partícula de reconocimiento de señal. La aparición de esta secuencia durante la elongación de la cadena polipeptídica constituye la primera etapa del direccionamiento proteico hacia el retículo endoplásmico.
A medida que el ribosoma traduce el ARN mensajero, la secuencia señal emerge progresivamente a través del túnel de salida de la subunidad ribosómica mayor. En ese momento es reconocida por la partícula de reconocimiento de señal, la cual se une simultáneamente a la secuencia señal y al propio ribosoma. Esta interacción produce una disminución temporal de la velocidad de elongación, evitando que la proteína continúe sintetizándose completamente en el citoplasma antes de alcanzar su destino intracelular. Este mecanismo asegura una coordinación precisa entre síntesis proteica y transporte intracelular.
El complejo formado por ribosoma, ARN mensajero, cadena polipeptídica naciente y partícula de reconocimiento de señal es transportado hacia la membrana del retículo endoplásmico, donde interactúa con receptores específicos localizados en la superficie citosólica de dicha membrana. Estos receptores actúan como sitios de anclaje molecular que permiten el posicionamiento exacto del ribosoma sobre complejos proteicos especializados denominados translocones.
El translocón principal del retículo endoplásmico está constituido por el complejo Sec61, una estructura multiproteica que forma un canal acuoso regulable a través de la membrana. Una vez que el ribosoma se acopla al translocón, la partícula de reconocimiento de señal es liberada y reciclada para participar en nuevos ciclos de direccionamiento proteico. Posteriormente, la traducción se reanuda y la energía derivada de la elongación ribosómica impulsa el desplazamiento progresivo de la cadena polipeptídica a través del canal transmembrana.
Este fenómeno recibe el nombre de translocación cotraduccional porque la síntesis de la proteína y su transporte hacia la luz del retículo endoplásmico ocurren de manera simultánea. A diferencia de otros mecanismos de transporte intracelular que se producen después de completada la síntesis proteica, en este caso la proteína comienza a ingresar al retículo mientras todavía está siendo ensamblada por el ribosoma. Esta estrategia minimiza el riesgo de plegamiento incorrecto en el citoplasma y favorece la adquisición temprana de su conformación tridimensional adecuada.
Conforme la cadena polipeptídica atraviesa el translocón, el lumen del retículo endoplásmico proporciona un entorno especializado para el procesamiento inicial de las proteínas. Este compartimento contiene numerosas proteínas auxiliares denominadas chaperonas moleculares, cuya función consiste en facilitar el plegamiento correcto de las cadenas polipeptídicas recién sintetizadas. Entre ellas destacan BiP, calnexina, calreticulina y otras proteínas residentes del retículo que participan activamente en el control de calidad proteico.
La importancia de este control de calidad radica en que una proteína incorrectamente plegada puede perder su función biológica, agregarse formando complejos tóxicos o desencadenar alteraciones celulares graves. Por esta razón, el retículo endoplásmico posee sistemas altamente sofisticados capaces de identificar proteínas defectuosas y dirigirlas hacia mecanismos de degradación antes de que alcancen otros compartimentos celulares.
Además del plegamiento, muchas proteínas experimentan modificaciones químicas iniciales durante su tránsito por la luz reticular. Una de las más importantes es la N-glicosilación, proceso mediante el cual cadenas de oligosacáridos preensambladas son transferidas a residuos específicos de asparagina presentes en la proteína naciente. Esta modificación influye en la estabilidad estructural, el reconocimiento molecular, el transporte intracelular y la función biológica final de numerosas proteínas secretadas y de membrana.
La formación de puentes disulfuro constituye otro acontecimiento esencial que ocurre dentro de la luz del retículo endoplásmico. El ambiente oxidante presente en este compartimento favorece la creación de enlaces covalentes entre residuos de cisteína, contribuyendo significativamente a la estabilidad tridimensional de muchas proteínas. Enzimas especializadas catalizan y corrigen la formación de estos enlaces hasta alcanzar la configuración adecuada.
La asociación masiva de ribosomas a la membrana del retículo endoplásmico produce cambios morfológicos fácilmente observables mediante microscopía electrónica. Las partículas ribosómicas adheridas a la superficie citosólica generan una apariencia rugosa o granular característica, motivo por el cual estas regiones reciben el nombre de retículo endoplásmico rugoso. Cada uno de los pequeños gránulos observados corresponde a un ribosoma individual o a grupos de ribosomas simultáneamente involucrados en la síntesis proteica.
Esta organización estructural refleja una extraordinaria eficiencia funcional. En células altamente secretoras, como las células acinares pancreáticas, los plasmocitos o determinadas células endocrinas, el retículo endoplásmico rugoso ocupa una proporción considerable del volumen citoplasmático. Ello permite sostener tasas elevadas de síntesis proteica requeridas para la producción continua de enzimas digestivas, anticuerpos, hormonas peptídicas y otras proteínas exportadas al medio extracelular.
Un aspecto particularmente importante es que la traducción raramente ocurre mediante un único ribosoma asociado a cada molécula de ARN mensajero. Habitualmente múltiples ribosomas se unen simultáneamente al mismo transcrito formando complejos conocidos como polirribosomas o polisomas. Esta organización incrementa de manera notable la eficiencia biosintética celular.
En un polisoma, diversos ribosomas avanzan secuencialmente a lo largo de una misma molécula de ARN mensajero, separados por distancias suficientes para evitar interferencias estéricas. Cada ribosoma sintetiza una copia independiente de la proteína codificada, de modo que una sola molécula de ARN mensajero puede generar numerosas cadenas polipeptídicas simultáneamente. Este mecanismo amplifica considerablemente la capacidad productiva de la célula sin necesidad de sintetizar múltiples copias adicionales del mismo ARN mensajero.
Cuando los polisomas se encuentran asociados al retículo endoplásmico, cada ribosoma individual permanece acoplado a su propio translocón mientras traduce la proteína correspondiente. Como resultado, múltiples cadenas polipeptídicas atraviesan simultáneamente la membrana reticular e ingresan a la luz del organelo. Las imágenes obtenidas mediante microscopía electrónica muestran con claridad esta disposición, revelando conjuntos de ribosomas alineados sobre una misma molécula de ARN mensajero y conectados funcionalmente con canales de translocación activos.
La existencia de numerosos polisomas asociados al retículo endoplásmico permite que una célula produzca grandes cantidades de proteínas secretoras o de membrana en períodos relativamente cortos. Este mecanismo resulta esencial para mantener procesos fisiológicos como la secreción hormonal, la producción de inmunoglobulinas, la síntesis de proteínas plasmáticas hepáticas, la renovación de receptores membranales y la formación de componentes extracelulares de los tejidos.
La coordinación entre reconocimiento de secuencias señal, reclutamiento ribosómico, acoplamiento al translocón, translocación cotraduccional, plegamiento proteico, modificaciones postraduccionales iniciales y organización en polisomas representa uno de los ejemplos más sofisticados de integración funcional dentro de la célula eucariótica. Gracias a este sistema, las proteínas son sintetizadas, clasificadas y distribuidas con una precisión extraordinaria, garantizando la organización estructural y funcional de prácticamente todos los compartimentos celulares.


Fuente y lecturas recomendadas:
- Blobel, G., & Dobberstein, B. (1975). Transfer of proteins across membranes. I. Presence of proteolytically processed and unprocessed nascent immunoglobulin light chains on membrane-bound ribosomes of murine myeloma. Journal of Cell Biology, 67(3), 835–851. https://doi.org/10.1083/jcb.67.3.835
- Blobel, G. (1980). Intracellular protein topogenesis. Proceedings of the National Academy of Sciences, 77(3), 1496–1500. https://doi.org/10.1073/pnas.77.3.1496
- Cross, B. C. S., Sinning, I., Luirink, J., & High, S. (2009). Delivering proteins for export from the cytosol. Nature Reviews Molecular Cell Biology, 10(4), 255–264. https://doi.org/10.1038/nrm2657
- Hegde, R. S., & Keenan, R. J. (2011). Tail-anchored membrane protein insertion into the endoplasmic reticulum. Nature Reviews Molecular Cell Biology, 12(12), 787–798. https://doi.org/10.1038/nrm3226
- Kellokumpu, S. (2019). Golgi pH, ion and redox homeostasis: How much do they really matter? Frontiers in Cell and Developmental Biology, 7, 93. https://doi.org/10.3389/fcell.2019.00093
- Park, E., & Rapoport, T. A. (2012). Mechanisms of Sec61/SecY-mediated protein translocation across membranes. Annual Review of Biophysics, 41, 21–40. https://doi.org/10.1146/annurev-biophys-050511-102312
- Schröder, M., & Kaufman, R. J. (2005). The mammalian unfolded protein response. Annual Review of Biochemistry, 74, 739–789. https://doi.org/10.1146/annurev.biochem.73.011303.074134
- Walter, P., & Blobel, G. (1981). Translocation of proteins across the endoplasmic reticulum. III. Signal recognition particle causes signal sequence-dependent and site-specific arrest of chain elongation that is released by microsomal membranes. Journal of Cell Biology, 91(2), 557–561. https://doi.org/10.1083/jcb.91.2.557
- Walter, P., & Johnson, A. E. (1994). Signal sequence recognition and protein targeting to the endoplasmic reticulum membrane. Annual Review of Cell Biology, 10, 87–119. https://doi.org/10.1146/annurev.cb.10.110194.000511
- Wickner, W., & Schekman, R. (2005). Protein translocation across biological membranes. Science, 310(5753), 1452–1456. https://doi.org/10.1126/science.1113752
- Zimmermann, R., Eyrisch, S., Ahmad, M., & Helms, V. (2011). Protein translocation across the endoplasmic reticulum membrane. Biochimica et Biophysica Acta, 1808(3), 912–924. https://doi.org/10.1016/j.bbamem.2010.06.015
Aprende administración paso a paso

ADMINISTRACION DESDE CERO