El movimiento amebiano constituye una forma especializada de locomoción celular mediante la cual una célula completa desplaza activamente su cuerpo a través de un medio líquido, una superficie biológica o una matriz extracelular. Este mecanismo recibe su nombre debido a que fue descrito inicialmente en amebas, organismos unicelulares que exhiben desplazamientos caracterizados por la emisión continua de prolongaciones citoplasmáticas denominadas seudópodos. Posteriormente se comprobó que numerosos tipos celulares de organismos multicelulares emplean principios biomecánicos similares para migrar durante procesos fisiológicos y patológicos fundamentales, incluyendo la respuesta inmunitaria, la reparación tisular, el desarrollo embrionario y la diseminación tumoral.
El movimiento amebiano representa uno de los ejemplos más complejos de integración entre señales bioquímicas, reorganización del citoesqueleto, adhesión celular, remodelación de membranas y producción de fuerza mecánica. Su estudio ha permitido comprender cómo las células son capaces de detectar estímulos ambientales, establecer una polaridad funcional, generar protrusiones dirigidas y transformar energía química en trabajo mecánico eficiente.
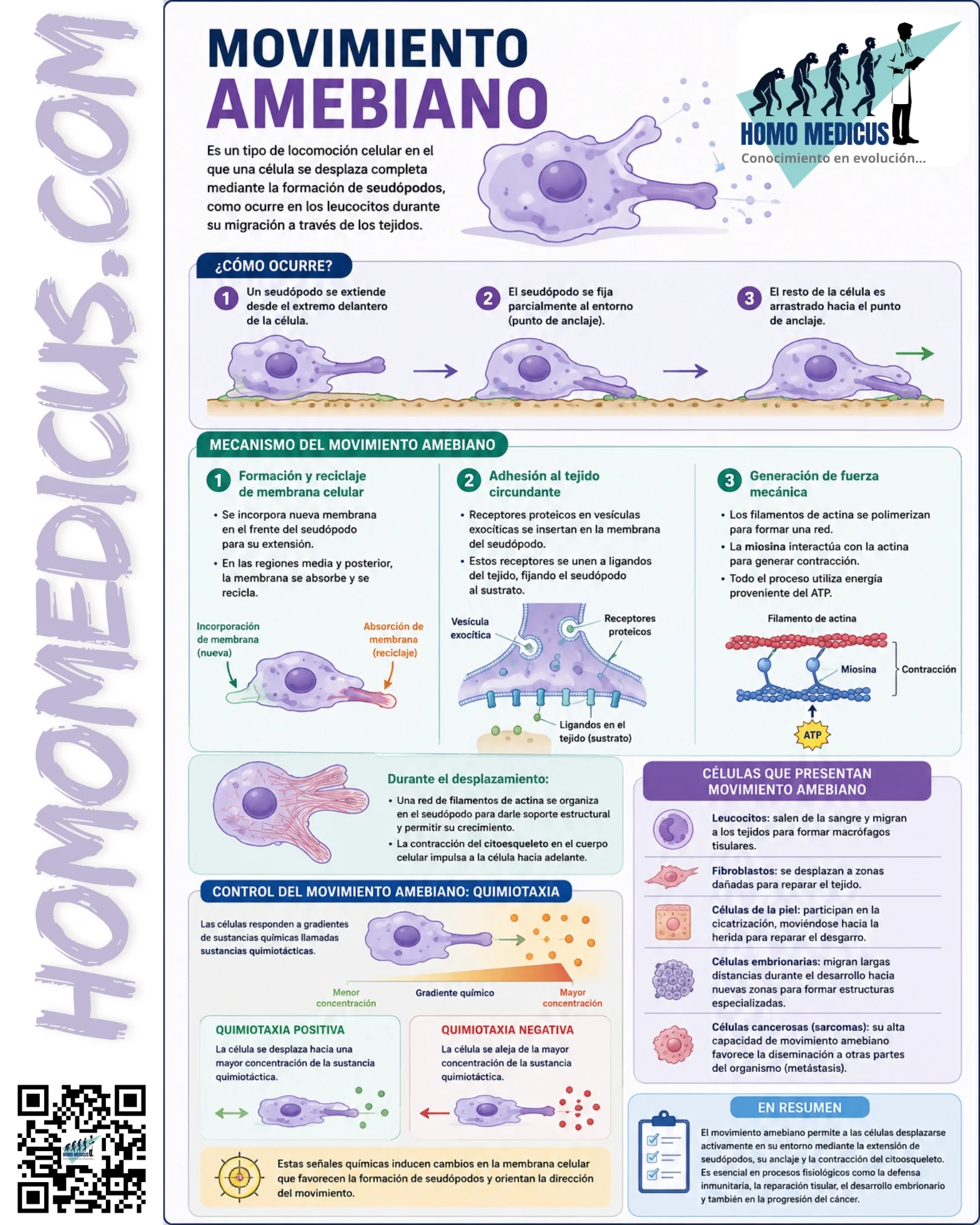
El desplazamiento amebiano implica el movimiento coordinado de toda la célula respecto de su entorno. A diferencia de otros mecanismos de locomoción celular, como el movimiento ciliar o flagelar, en el movimiento amebiano no existe una estructura locomotora permanente. En cambio, la célula modifica constantemente su forma mediante reorganizaciones dinámicas del citoplasma y del citoesqueleto.
La locomoción amebiana puede dividirse en varias etapas funcionales interdependientes:
- Polarización celular.
- Formación del seudópodo.
- Adhesión al sustrato.
- Generación de fuerza contráctil.
- Retracción de la región posterior.
- Reciclaje de membrana y componentes celulares.
Cada una de estas etapas depende de complejas redes de señalización molecular y de la participación coordinada de numerosas proteínas reguladoras.
Antes de desplazarse, la célula debe adquirir una polaridad estructural definida. Esto significa que una región celular se especializa como extremo anterior o director, mientras que otra región se convierte en extremo posterior.
La polarización celular ocurre en respuesta a estímulos externos, especialmente gradientes químicos, señales mecánicas o interacciones con la matriz extracelular. La distribución asimétrica de receptores de membrana y moléculas señalizadoras provoca una reorganización diferencial del citoesqueleto, generando una región frontal rica en polimerización de actina y una región posterior enriquecida en proteínas contráctiles asociadas con miosina II.
Esta polarización constituye el requisito esencial para que la célula pueda distinguir una dirección de avance y mantener una migración persistente.
La característica más evidente del movimiento amebiano es la formación de seudópodos. Los seudópodos son extensiones temporales del citoplasma que protruyen desde la superficie celular en dirección al desplazamiento. Su formación depende principalmente de la rápida polimerización de filamentos de actina debajo de la membrana plasmática.
La proteína actina existe inicialmente en forma de monómeros solubles. Cuando la célula recibe señales apropiadas, estos monómeros se ensamblan formando filamentos que empujan mecánicamente la membrana plasmática hacia el exterior. Como resultado aparece una protrusión que se extiende progresivamente sobre el sustrato.
La generación de estas estructuras requiere la participación de complejos moleculares especializados que regulan la nucleación, elongación y ramificación de los filamentos de actina. La formación de redes tridimensionales densamente ramificadas proporciona soporte mecánico suficiente para impulsar el crecimiento del seudópodo.
El crecimiento continuo de la protrusión determina la dirección inmediata del desplazamiento celular. Durante muchos años se propuso que el movimiento amebiano dependía principalmente de la incorporación de nueva membrana en la región anterior y de la absorción de membrana en la región posterior.
Actualmente se sabe que el reciclaje de membrana efectivamente participa en la migración celular, aunque no constituye el único mecanismo impulsor. Las células migratorias mantienen un intenso tráfico vesicular mediante procesos de exocitosis y endocitosis.
Las vesículas intracelulares transportan proteínas de adhesión, receptores de membrana y lípidos hacia el frente migratorio. Simultáneamente, regiones de membrana localizadas en zonas posteriores son internalizadas y recicladas.
Este flujo continuo contribuye a mantener la superficie celular necesaria para la extensión del seudópodo y favorece la redistribución de moléculas esenciales para la locomoción.
La protrusión aislada del seudópodo no es suficiente para producir movimiento efectivo. Para que la célula avance resulta indispensable que el extremo anterior establezca puntos de anclaje transitorios con el entorno. Estas adhesiones son mediadas principalmente por receptores transmembrana especializados denominados integrinas. Las integrinas reconocen componentes de la matriz extracelular, incluyendo fibronectina, laminina, colágeno y vitronectina.
Cuando el seudópodo entra en contacto con una superficie adecuada, las integrinas se agrupan formando complejos de adhesión. Dichos complejos conectan mecánicamente el exterior celular con la red intracelular de filamentos de actina. La formación de adhesiones focales permite que las fuerzas generadas internamente se transmitan al sustrato. De esta manera, el seudópodo queda temporalmente fijado mientras el resto de la célula es impulsado hacia adelante.
Las adhesiones celulares son altamente dinámicas. Nuevos puntos de anclaje se forman continuamente en el frente migratorio, mientras que los existentes en la región posterior son desmantelados para permitir el avance.
La locomoción celular requiere energía y sistemas capaces de transformar dicha energía en movimiento. El principal motor molecular del movimiento amebiano es la interacción entre actina y miosina.
La actina constituye el componente estructural fundamental del citoesqueleto migratorio. La miosina II actúa como proteína motora capaz de deslizar filamentos de actina unos respecto de otros.
Cuando las moléculas de miosina interactúan con la red de actina, generan tensiones contráctiles que producen acortamiento local del citoesqueleto. Estas contracciones ayudan a desplazar el cuerpo celular hacia el punto de anclaje establecido por el seudópodo.
La energía necesaria para este proceso proviene de la hidrólisis de ATP. Cada ciclo mecánico de la miosina implica unión a actina, generación de fuerza, liberación y reinicio del ciclo mediante consumo de ATP. Por tanto, la locomoción amebiana representa un ejemplo directo de conversión de energía química en energía mecánica.
El citoesqueleto de actina constituye el elemento central de todo el movimiento amebiano. En la región anterior predominan procesos de ensamblaje de filamentos. Esta polimerización impulsa la expansión del seudópodo. En la región media se generan estructuras de soporte mecánico que estabilizan la forma celular. En la región posterior predominan procesos contráctiles mediados por miosina II que facilitan la retracción del extremo caudal.
La coordinación espacial de estos fenómenos permite que la célula mantenga una migración eficiente y direccional. La desorganización experimental de los filamentos de actina bloquea prácticamente toda forma de movimiento amebiano, demostrando la importancia crítica de este sistema.
Mientras el frente celular se extiende y se adhiere al sustrato, la región posterior debe retraerse para completar el desplazamiento. Esta retracción depende de la contracción actomiosínica y de la desintegración de las adhesiones existentes en la parte posterior de la célula. La tensión generada por la miosina provoca el colapso progresivo de la cola celular, permitiendo que el citoplasma fluya hacia adelante. Como resultado, la masa celular completa se desplaza en dirección al seudópodo previamente formado.
Un fenómeno clásico observado en células ameboides es el flujo citoplasmático interno. Durante la migración se produce una redistribución continua de componentes citoplasmáticos desde regiones posteriores hacia regiones anteriores. Este flujo contribuye al crecimiento del seudópodo y facilita la reorganización rápida de estructuras intracelulares necesarias para el desplazamiento.
Los cambios en las propiedades mecánicas del citoplasma permiten transiciones entre estados más fluidos y más gelificados, fenómeno que participa activamente en la locomoción celular.
Células humanas que presentan movimiento amebiano
Leucocitos
Los leucocitos constituyen el ejemplo fisiológico más representativo. Durante una respuesta inflamatoria, estas células abandonan la circulación sanguínea mediante extravasación vascular y migran a través de los tejidos hasta alcanzar el foco de infección o lesión.
Los neutrófilos pueden desplazarse rápidamente siguiendo gradientes de moléculas liberadas por bacterias o tejidos dañados. Los monocitos también migran mediante mecanismos ameboides antes de diferenciarse en macrófagos tisulares. La eficacia de esta migración resulta esencial para la defensa inmunitaria.
Fibroblastos
Los fibroblastos participan activamente en la cicatrización de heridas. Tras una lesión tisular migran hacia la zona afectada, donde sintetizan colágeno, fibronectina y otros componentes de la matriz extracelular. Su capacidad migratoria permite reconstruir la arquitectura tisular dañada y restablecer la integridad mecánica del tejido.
Células epiteliales
Durante la reparación de heridas cutáneas, las células epiteliales migran colectivamente para cubrir áreas lesionadas. Este proceso resulta indispensable para restaurar la barrera protectora de la piel.
Células embrionarias
Durante el desarrollo embrionario ocurren migraciones celulares masivas. Las células de la cresta neural, por ejemplo, recorren grandes distancias y originan numerosos tejidos especializados, incluyendo neuronas periféricas, melanocitos y estructuras craneofaciales. La correcta regulación del movimiento celular es indispensable para el desarrollo normal del organismo.
Movimiento amebiano y cáncer
Muchas células tumorales adquieren capacidades migratorias semejantes a las observadas en células normales durante el desarrollo embrionario.
Las células cancerosas pueden modificar dinámicamente su citoesqueleto, generar seudópodos y desplazarse a través de matrices extracelulares complejas. En numerosos tumores se observa una transición hacia modos de migración ameboide caracterizados por elevada plasticidad morfológica y capacidad para atravesar tejidos sin necesidad de degradar extensamente la matriz extracelular. Esta propiedad favorece la invasión local y la formación de metástasis.
Los sarcomas, debido a su origen mesenquimatoso, suelen presentar una notable capacidad migratoria basada en mecanismos ameboides.
Control del movimiento amebiano mediante quimiotaxia
La quimiotaxia constituye uno de los mecanismos más importantes para orientar el movimiento amebiano. Este fenómeno ocurre cuando una célula detecta diferencias espaciales en la concentración de determinadas moléculas presentes en el entorno.
Las sustancias capaces de inducir respuestas migratorias reciben el nombre de sustancias quimiotácticas. Los receptores localizados en la membrana celular detectan estas moléculas y activan cascadas intracelulares de señalización. Como consecuencia, la polimerización de actina aumenta en el lado de la célula más expuesto al estímulo. La formación preferencial de seudópodos en esa región orienta el desplazamiento hacia la fuente de la señal química.
Quimiotaxia positiva: La quimiotaxia positiva ocurre cuando la célula se desplaza hacia concentraciones crecientes del estímulo químico. Este mecanismo permite que neutrófilos, monocitos y otras células inmunitarias localicen rápidamente focos infecciosos o áreas de daño tisular.
Quimiotaxia negativa: La quimiotaxia negativa ocurre cuando la célula se aleja de la fuente de la sustancia química. Aunque es menos frecuente que la quimiotaxia positiva, desempeña funciones importantes en determinados procesos de desarrollo y organización tisular.
La región de la membrana celular que recibe la mayor intensidad de estimulación química experimenta una reorganización molecular profunda. Se activan proteínas reguladoras de la actina, aumenta la formación de fosfolípidos señalizadores y se produce una acumulación local de complejos responsables de la polimerización citosquelética. Como consecuencia surge una protrusión dirigida que se convierte en el nuevo frente migratorio de la célula.
Este mecanismo permite transformar información química proveniente del ambiente en movimiento orientado y constituye uno de los ejemplos más sofisticados de transducción de señales biológicas.
El movimiento amebiano es un proceso biológico altamente dinámico que depende de la integración coordinada entre polarización celular, formación de seudópodos, adhesión al sustrato, reorganización del citoesqueleto de actina, actividad de miosina, consumo de ATP y respuestas quimiotácticas. Gracias a estos mecanismos, las células pueden desplazarse de forma dirigida a través de tejidos complejos, permitiendo funciones esenciales como la inmunidad, la cicatrización, el desarrollo embrionario y, en condiciones patológicas, la invasión tumoral y la metástasis.

Fuente y lecturas recomendadas:
- Abercrombie, M. (1980). The crawling movement of metazoan cells. Proceedings of the Royal Society of London. Series B, Biological Sciences, 207(1167), 129–147. https://doi.org/10.1098/rspb.1980.0017
- Charras, G., & Sahai, E. (2014). Physical influences of the extracellular environment on cell migration. Nature Reviews Molecular Cell Biology, 15(12), 813–824. https://doi.org/10.1038/nrm3897
- Friedl, P., & Wolf, K. (2003). Tumour-cell invasion and migration: Diversity and escape mechanisms. Nature Reviews Cancer, 3(5), 362–374. https://doi.org/10.1038/nrc1075
- Friedl, P., & Wolf, K. (2010). Plasticity of cell migration: A multiscale tuning model. The Journal of Cell Biology, 188(1), 11–19. https://doi.org/10.1083/jcb.200909003
- Insall, R. H., & Machesky, L. M. (2009). Actin dynamics at the leading edge: From simple machinery to complex networks. Developmental Cell, 17(3), 310–322. https://doi.org/10.1016/j.devcel.2009.08.012
- Lauffenburger, D. A., & Horwitz, A. F. (1996). Cell migration: A physically integrated molecular process. Cell, 84(3), 359–369. https://doi.org/10.1016/S0092-8674(00)81280-5
- Ridley, A. J., Schwartz, M. A., Burridge, K., Firtel, R. A., Ginsberg, M. H., Borisy, G., Parsons, J. T., & Horwitz, A. R. (2003). Cell migration: Integrating signals from front to back. Science, 302(5651), 1704–1709. https://doi.org/10.1126/science.1092053
- Rorth, P. (2009). Collective cell migration. Annual Review of Cell and Developmental Biology, 25, 407–429. https://doi.org/10.1146/annurev.cellbio.042308.113231
- Sheetz, M. P., Felsenfeld, D., & Galbraith, C. G. (1998). Cell migration: Regulation of force on extracellular-matrix-integrin complexes. Trends in Cell Biology, 8(2), 51–54. https://doi.org/10.1016/S0962-8924(97)01147-6
- Van Haastert, P. J. M., & Devreotes, P. N. (2004). Chemotaxis: Signalling the way forward. Nature Reviews Molecular Cell Biology, 5(8), 626–634. https://doi.org/10.1038/nrm1435
- Yamada, K. M., & Sixt, M. (2019). Mechanisms of 3D cell migration. Nature Reviews Molecular Cell Biology, 20(12), 738–752. https://doi.org/10.1038/s41580-019-0172-9
Aprende administración paso a paso

ADMINISTRACION DESDE CERO