El movimiento ciliar constituye uno de los ejemplos más sofisticados de conversión biológica de energía química en trabajo mecánico. Este proceso permite el desplazamiento de fluidos, partículas o células completas mediante la generación de ondas de flexión altamente coordinadas a lo largo del cilio. Aunque numerosos aspectos de su regulación fina continúan siendo objeto de investigación, los principios fundamentales que explican la producción del movimiento han sido establecidos mediante estudios estructurales, bioquímicos, genéticos y biofísicos realizados durante varias décadas.
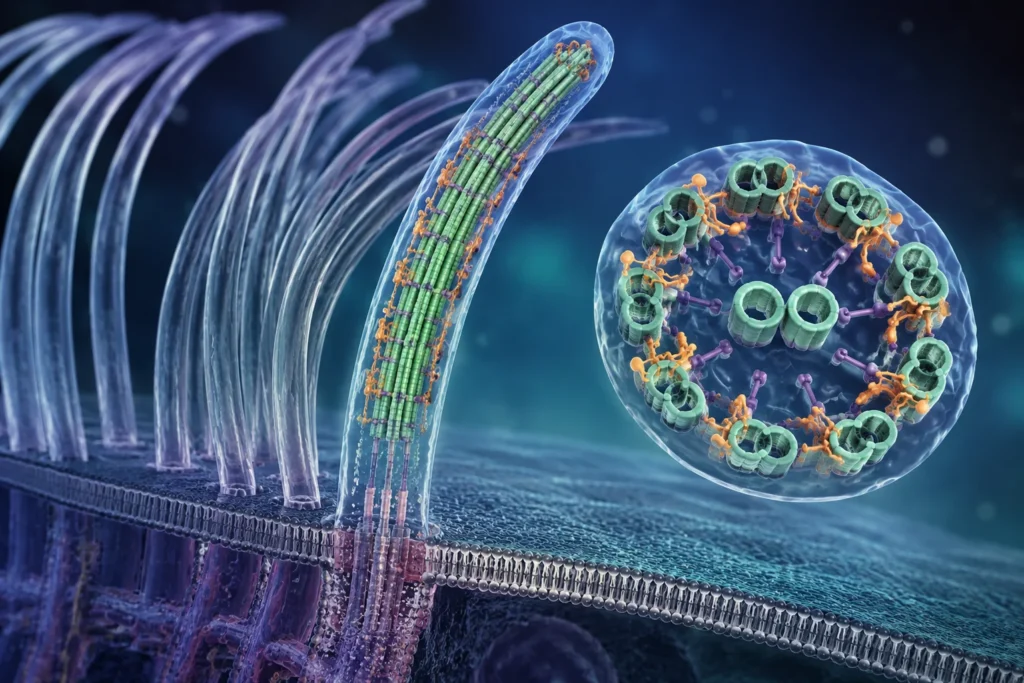
La estructura responsable del movimiento recibe el nombre de axonema, que constituye el armazón interno del cilio. En los cilios móviles típicos, el axonema presenta la organización denominada 9 + 2, formada por nueve dobletes periféricos de microtúbulos que rodean un par central de microtúbulos sencillos. Esta disposición no representa únicamente un soporte estructural, sino una máquina molecular compleja integrada por centenares de proteínas especializadas. Los dobletes periféricos se encuentran interconectados mediante enlaces proteicos, entre los que destacan los enlaces de nexina y los radios radiales, que mantienen la integridad mecánica del conjunto y permiten la coordinación funcional entre los distintos componentes.
La importancia central del axonema quedó demostrada cuando diversos experimentos mostraron que los cilios podían continuar moviéndose incluso después de eliminar la membrana plasmática y gran parte de los componentes celulares asociados. Cuando se preserva el axonema y se suministran condiciones químicas adecuadas, el batido ciliar puede reactivarse artificialmente. Este hallazgo demostró que la maquinaria esencial para la generación del movimiento reside dentro del propio axonema y no depende directamente de otras estructuras celulares para producir cada ciclo de flexión.
Uno de los requisitos fundamentales para que el movimiento continúe es la disponibilidad de ATP, que constituye la fuente inmediata de energía para los motores moleculares axonemales. El segundo requisito es la existencia de condiciones iónicas apropiadas, particularmente concentraciones adecuadas de Mg²⁺ y Ca²⁺. El magnesio participa directamente en la utilización del ATP por las proteínas motoras, mientras que el calcio actúa como un regulador importante de la frecuencia, amplitud y patrón del batido ciliar. Las modificaciones en la concentración intracelular de calcio pueden transformar de manera significativa la forma de las ondas de flexión producidas por el axonema.
El elemento generador de fuerza dentro del axonema está constituido por los brazos de dineína, complejos proteicos motores anclados sobre los microtúbulos periféricos. Cada brazo de dineína se proyecta desde el túbulo A de un doblete hacia el túbulo B del doblete vecino. Estas proteínas poseen actividad ATPasa, lo que significa que pueden hidrolizar ATP y transformar la energía química liberada en movimiento mecánico.
Cuando una molécula de ATP se une a la dineína y posteriormente es hidrolizada, la proteína experimenta cambios conformacionales que generan desplazamientos dirigidos a lo largo del microtúbulo adyacente. Las cabezas motoras de la dineína realizan ciclos repetidos de unión, generación de fuerza y liberación, produciendo un fenómeno comparable al desplazamiento progresivo de una estructura sobre otra. Como resultado, un doblete microtubular intenta deslizarse respecto al doblete vecino.
Sin embargo, si los dobletes pudieran deslizarse libremente unos respecto a otros, el axonema simplemente se alargaría o desorganizaría sin producir una curvatura efectiva. Esto no ocurre debido a la presencia de los enlaces de nexina, los radios radiales y otros complejos estructurales que restringen el deslizamiento excesivo. Como consecuencia, la fuerza generada por la dineína no se traduce en un desplazamiento lineal libre, sino en una deformación mecánica del conjunto. El deslizamiento relativo entre dobletes se convierte así en una curvatura del axonema.
Durante una fase de flexión determinada, los dobletes localizados en un lado del axonema desarrollan actividad motora intensa, mientras que los situados en el lado opuesto permanecen relativamente inactivos. Los dobletes activos ejercen fuerzas que provocan el desplazamiento relativo de los microtúbulos y generan una curvatura en una dirección concreta. Posteriormente, la actividad motora cambia de lado y la curvatura se produce en sentido contrario. La alternancia espacial y temporal de la actividad de la dineína permite la generación de ondas repetitivas de flexión que se propagan a lo largo del cilio.
El denominado movimiento anterógrado del cilio puede entenderse precisamente mediante este mecanismo. Los dobletes localizados en el borde frontal del cilio experimentan un desplazamiento relativo hacia la punta, mientras que los situados en el borde posterior permanecen comparativamente estacionarios. La diferencia de desplazamiento entre ambas regiones produce una deformación mecánica que se manifiesta como una flexión dirigida del axonema. La repetición secuencial de este fenómeno genera la onda característica de batido observada en los cilios móviles.
Aunque el mecanismo básico de generación de fuerza está bien establecido, la forma exacta en que se coordina la activación de miles de moléculas de dineína sigue siendo una de las cuestiones más complejas de la biología ciliar. Se sabe que los motores no funcionan de manera independiente; por el contrario, deben activarse y desactivarse de forma sincronizada para producir un patrón ordenado de movimiento. Diversas evidencias indican que esta coordinación depende de un sistema regulador integrado por el par central de microtúbulos, los radios radiales y los complejos reguladores asociados a la dineína.
Los radios radiales son estructuras proteicas que se extienden desde cada doblete periférico hacia el par central. Estas estructuras funcionan como transmisores mecanoquímicos capaces de comunicar información reguladora entre el aparato central y los motores de dineína. Su organización tridimensional y su composición proteica sugieren que participan en la modulación precisa de la actividad motora necesaria para producir patrones normales de batido.
El par central de microtúbulos posee una relevancia especial porque numerosos estudios han demostrado que los cilios que carecen de esta estructura presentan graves alteraciones de motilidad o son completamente inmóviles. La evidencia acumulada indica que el aparato central actúa como un centro regulador que distribuye señales hacia los radios radiales y, a través de ellos, hacia los brazos de dineína. Estas señales pueden involucrar mecanismos mecánicos, bioquímicos y electroquímicos que controlan qué grupos de motores deben activarse en cada instante.
Además de la regulación estructural, existen mecanismos bioquímicos complejos basados en proteínas cinasas, fosfatasas, calmodulina y otras moléculas señalizadoras localizadas dentro del axonema. Estas proteínas modifican el estado funcional de los motores de dineína mediante procesos de fosforilación y desfosforilación, ajustando la intensidad y sincronización de las fuerzas generadas. De esta manera, el axonema no funciona como una simple estructura mecánica, sino como un sistema integrado de señalización y generación de fuerza.
El calcio desempeña asimismo una función reguladora fundamental. Los cambios en la concentración intraciliar de este ion modifican la actividad de diversas proteínas asociadas al axonema y alteran el patrón de activación de la dineína. Gracias a este mecanismo, los cilios pueden adaptar su frecuencia de batido, la amplitud de sus ondas y la dirección de sus movimientos en respuesta a diferentes estímulos fisiológicos.
El movimiento ciliar surge de la interacción coordinada entre la hidrólisis de ATP por los brazos de dineína, el deslizamiento relativo de los dobletes microtubulares, las restricciones mecánicas impuestas por los enlaces estructurales del axonema y los sistemas reguladores asociados al par central y los radios radiales. La energía química almacenada en el ATP se transforma en fuerzas de deslizamiento; estas fuerzas se convierten en curvaturas debido a las restricciones estructurales del axonema; y la secuencia espacial y temporal de activación de los motores genera finalmente las ondas de flexión que caracterizan el batido ciliar. Aunque muchos detalles moleculares continúan investigándose, la evidencia disponible demuestra que el cilio constituye una de las máquinas biológicas más complejas y eficientes conocidas, capaz de coordinar miles de motores moleculares para producir un movimiento altamente ordenado y repetitivo.


Fuente y lecturas recomendadas:
- King, S. M. (2017). Ciliary motility: Regulation of axonemal dynein motors. Cold Spring Harbor Perspectives in Biology, 10(1), a018325. https://doi.org/10.1101/cshperspect.a018325
- Lindemann, C. B. (2024). The mechanics of cilia and flagella: What we know and what we need to know. Cytoskeleton, 81(1), 3–22. https://doi.org/10.1002/cm.21879
- Mitchell, D. R. (2010). Flagellar and ciliary beating: The proven and the possible. Journal of Cell Science, 123(4), 519–528. https://doi.org/10.1242/jcs.051326
- Pigino, G., & Ishikawa, T. (2012). Axonemal radial spokes: 3D structure, function and assembly. Bioarchitecture, 2(2), 50–58. https://doi.org/10.4161/bioa.20394
- Porter, M. E., & Sale, W. S. (2000). The 9 + 2 axoneme anchors multiple inner arm dyneins and a network of regulatory complexes that control ciliary motility. Journal of Cell Biology, 151(5), F37–F42.
- Satir, P., & Christensen, S. T. (2007). Overview of structure and function of mammalian cilia. Annual Review of Physiology, 69, 377–400. https://doi.org/10.1146/annurev.physiol.69.040705.141236
- Smith, E. F., & Yang, P. (2004). The radial spokes and central apparatus: Mechano-chemical transducers that regulate flagellar motility. Cell Motility and the Cytoskeleton, 57(1), 8–17. https://doi.org/10.1002/cm.10155
- Wirschell, M., Yang, C., Yang, P., Fox, L., Yanagisawa, H. A., Kamiya, R., Witman, G. B., Porter, M. E., & Sale, W. S. (2008). Building a radial spoke: Flagellar radial spoke protein 3 forms a scaffold for assembly of the radial spoke complex. Molecular Biology of the Cell, 19(7), 2912–2921. https://doi.org/10.1091/mbc.e08-01-0064
- Ziętkiewicz, E., Bukowy-Bieryłło, Z., & Voelkel, K. (2021). Central apparatus, the molecular kickstarter of ciliary and flagellar nanomachines. International Journal of Molecular Sciences, 22(6), 3013. https://doi.org/10.3390/ijms22063013
Aprende administración paso a paso

ADMINISTRACION DESDE CERO