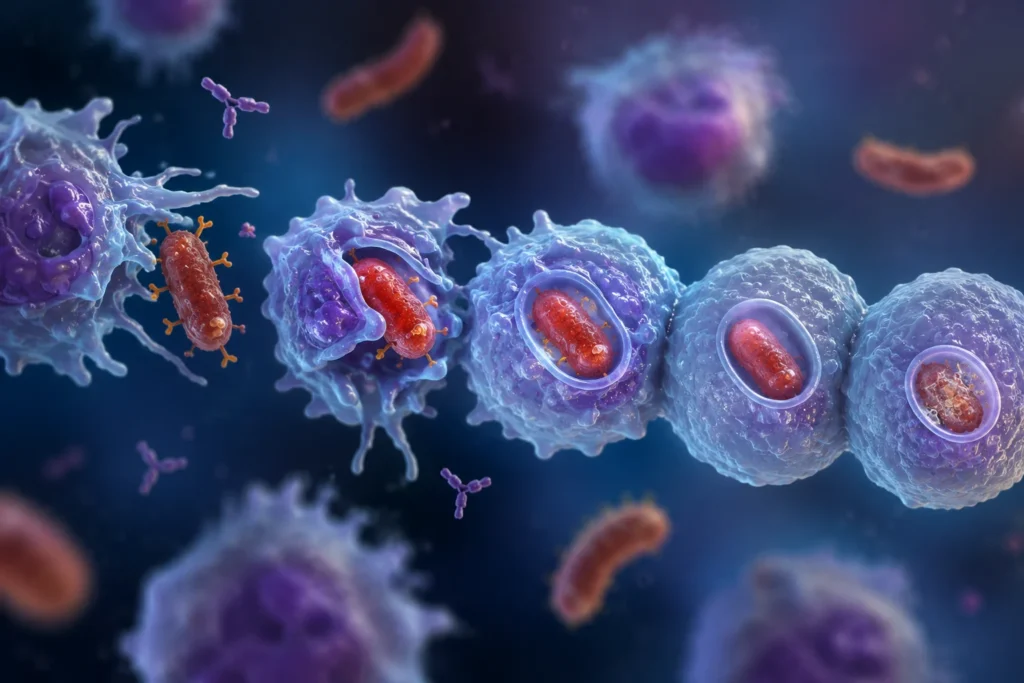
La fagocitosis es un proceso biológico altamente especializado mediante el cual determinadas células del sistema inmunitario reconocen, rodean, internalizan y destruyen partículas sólidas de gran tamaño, incluyendo bacterias, hongos, células envejecidas, células apoptóticas, restos tisulares y diversos cuerpos extraños. A diferencia de la pinocitosis, que incorpora principalmente líquidos y moléculas disueltas del medio extracelular, la fagocitosis está diseñada para la captación de estructuras cuyo tamaño supera ampliamente al de las macromoléculas individuales y que, por tanto, requieren una reorganización profunda de la membrana plasmática y del citoesqueleto celular para ser incorporadas al interior de la célula. Esta función constituye uno de los mecanismos fundamentales de la inmunidad innata y representa una de las estrategias más antiguas y conservadas evolutivamente para la defensa frente a microorganismos y para el mantenimiento de la homeostasis tisular.
Solo ciertos tipos celulares poseen una capacidad fagocítica plenamente desarrollada. Entre ellos destacan los macrófagos tisulares, los monocitos circulantes, los neutrófilos, algunas células dendríticas y otros fagocitos profesionales. Estas células expresan en su superficie una amplia variedad de receptores especializados capaces de identificar microorganismos, restos celulares y moléculas que indican daño tisular. La presencia de estos receptores permite que la fagocitosis no sea un simple proceso mecánico de ingestión, sino un fenómeno altamente selectivo y regulado que integra señales procedentes del entorno extracelular para decidir qué partículas deben ser eliminadas y cómo debe responder posteriormente la célula.
La importancia fisiológica de la fagocitosis va mucho más allá de la eliminación física de partículas. La formación del fagosoma constituye un centro intracelular de procesamiento de información inmunológica. Durante la internalización, los receptores activados desencadenan complejas cascadas de señalización que permiten a la célula identificar la naturaleza del material ingerido, activar programas antimicrobianos específicos, producir citocinas inflamatorias y presentar antígenos a otras células del sistema inmunitario. En consecuencia, la fagocitosis participa simultáneamente en la defensa contra infecciones, en la eliminación de células dañadas y en la coordinación de respuestas inmunitarias adaptativas.
La fagocitosis comienza cuando una partícula entra en contacto con la superficie de un fagocito y establece interacciones con receptores específicos presentes en la membrana plasmática. Estos receptores reconocen moléculas denominadas ligandos, localizadas sobre la superficie de la partícula que va a ser ingerida. La unión receptor-ligando constituye el acontecimiento que inicia todo el proceso de internalización.
Los fagocitos expresan numerosos tipos de receptores capaces de reconocer diferentes clases de señales biológicas. Algunos identifican directamente componentes microbianos conservados, mientras que otros reconocen moléculas producidas durante el daño tisular o la muerte celular. Sin embargo, uno de los mecanismos más eficientes de reconocimiento ocurre cuando la partícula ha sido previamente marcada por moléculas del sistema inmunitario, fenómeno conocido como opsonización.
La opsonización consiste en el recubrimiento de una partícula mediante moléculas que facilitan su reconocimiento por los fagocitos. Entre las opsoninas más importantes se encuentran los anticuerpos y diversas proteínas del sistema del complemento. Cuando una bacteria queda cubierta por inmunoglobulinas específicas, las regiones Fc de dichos anticuerpos permanecen expuestas hacia el exterior. Los receptores Fc presentes en la superficie de los macrófagos y neutrófilos reconocen estas regiones y se unen a ellas, estableciendo una conexión firme entre la célula fagocítica y el microorganismo.
De manera similar, proteínas del complemento depositadas sobre la superficie microbiana pueden ser reconocidas por receptores específicos del complemento expresados por los fagocitos. Como resultado, la partícula se vuelve mucho más visible para el sistema inmunitario y la probabilidad de que sea capturada aumenta significativamente. Este mecanismo explica por qué microorganismos recubiertos por anticuerpos o componentes del complemento son eliminados con mucha mayor eficacia que aquellos que carecen de dichas marcas.
La opsonización posee una enorme importancia biológica porque incrementa la fuerza de adhesión entre la partícula y el fagocito. Además, la agrupación simultánea de numerosos receptores sobre la superficie celular amplifica las señales intracelulares que activan la reorganización del citoesqueleto y favorecen la formación del fagosoma.
Una vez que los ligandos de la partícula interactúan con los receptores correspondientes, comienza la primera etapa visible de la fagocitosis. Los receptores se concentran progresivamente en la zona de contacto y forman complejos moleculares altamente organizados. Esta agrupación de receptores desencadena cascadas de señalización intracelular que activan proteínas reguladoras, pequeñas GTPasas y numerosas moléculas responsables del remodelado del citoesqueleto.
La unión inicial no es un acontecimiento estático. Conforme aparecen nuevos contactos entre receptores y ligandos, la membrana continúa adhiriéndose a la superficie de la partícula. Este proceso genera una adhesión progresiva y altamente coordinada que permite a la célula comenzar a rodear el objeto que será ingerido.
Tras la adhesión inicial, la membrana plasmática empieza a extenderse alrededor de la partícula. Esta extensión no ocurre de forma aleatoria, sino mediante un mecanismo denominado mecanismo de cremallera. En este modelo, cada nueva unión entre receptores y ligandos favorece la formación de un contacto adicional inmediatamente adyacente, de manera semejante al cierre progresivo de una cremallera.
La membrana celular genera prolongaciones que avanzan sobre la superficie de la partícula mientras nuevas moléculas receptoras se unen continuamente a nuevos ligandos. Como consecuencia, la célula va envolviendo progresivamente el objeto hasta cubrirlo por completo. Este proceso requiere una extraordinaria reorganización estructural de la membrana y del citoesqueleto subyacente.
La formación de la llamada copa fagocítica constituye una de las manifestaciones más características de la fagocitosis. Esta estructura semiesférica rodea gradualmente la partícula mientras se produce una intensa polimerización de filamentos de actina en el borde de avance. Los filamentos recién formados generan fuerzas mecánicas que empujan la membrana hacia delante y permiten que la célula envuelva partículas cuyo tamaño puede superar varias veces el diámetro de una bacteria.
La actina desempeña un papel central durante toda la fagocitosis. La unión de los receptores activa complejas vías de señalización que inducen la polimerización rápida de filamentos de actina debajo de la membrana plasmática. Estos filamentos forman una red dinámica capaz de generar fuerzas mecánicas dirigidas.
La reorganización de la actina permite que la membrana se deforme, produzca prolongaciones y rodee completamente la partícula. Además, otros componentes del citoesqueleto colaboran estabilizando la estructura en formación y coordinando los movimientos necesarios para completar la internalización. La participación de la actina explica por qué la fagocitosis es un proceso activo que consume energía metabólica y depende del funcionamiento normal de múltiples proteínas contráctiles y reguladoras.
Cuando la membrana ha rodeado prácticamente toda la partícula, la copa fagocítica se transforma progresivamente en una vesícula cerrada. Durante esta etapa, las proteínas del citoesqueleto continúan generando fuerzas que impulsan la estructura hacia el interior del citoplasma. La actina y diversas proteínas contráctiles forman complejos alrededor de la vesícula naciente y contribuyen a su desplazamiento intracelular.
Este movimiento asegura que el material ingerido abandone la superficie celular y sea transportado hacia regiones internas donde podrá iniciar las etapas posteriores de procesamiento y destrucción. La coordinación entre membrana, citoesqueleto y proteínas motoras permite que la internalización se complete en un tiempo relativamente corto pese al gran tamaño de muchas partículas fagocitadas.
La fase final de la internalización ocurre cuando los bordes de la copa fagocítica se encuentran y forman un estrecho cuello membranoso que conecta la vesícula con la superficie celular. Diversas proteínas contráctiles ejercen presión sobre esta región hasta provocar su estrangulamiento y posterior separación. Como resultado, la vesícula queda completamente aislada dentro del citoplasma.
La estructura recién formada recibe el nombre de fagosoma. Este orgánulo contiene en su interior la partícula capturada y constituye el punto de partida para una serie de transformaciones posteriores que culminarán con la destrucción o procesamiento del material ingerido.
La creación del fagosoma no representa el final de la fagocitosis, sino el inicio de una nueva etapa funcional. Después de su formación, el fagosoma experimenta un proceso progresivo de maduración durante el cual interactúa con distintos compartimentos intracelulares. Estas interacciones modifican gradualmente su composición molecular y aumentan su capacidad microbicida.
A medida que madura, el fagosoma adquiere enzimas hidrolíticas, bombas de protones y múltiples proteínas especializadas que acidifican su interior y favorecen la degradación del material capturado. Finalmente, puede fusionarse con lisosomas para originar el fagolisosoma, una estructura altamente especializada en la destrucción de microorganismos y restos celulares.
Durante este proceso también se generan especies reactivas de oxígeno y otras moléculas antimicrobianas que contribuyen a la eliminación eficiente de patógenos. Simultáneamente, los productos resultantes de la degradación pueden ser utilizados para la presentación antigénica y para la activación de respuestas inmunitarias posteriores.
La fagocitosis es esencial para la supervivencia de los organismos multicelulares. Gracias a ella se eliminan microorganismos invasores antes de que puedan multiplicarse y producir enfermedad. Asimismo, permite retirar células envejecidas, células apoptóticas y restos de tejidos lesionados, evitando la acumulación de material potencialmente tóxico o inflamatorio.
Además de su función defensiva y de limpieza, la fagocitosis participa activamente en la regulación de la respuesta inmunitaria. La información obtenida durante el reconocimiento y procesamiento de las partículas ingeridas determina la producción de mediadores inflamatorios, la activación de otras células inmunitarias y la generación de respuestas adaptativas específicas. Por ello, la fagocitosis constituye simultáneamente un mecanismo de eliminación física, una plataforma de señalización celular y un sistema de adquisición de información inmunológica.


Fuente y lecturas recomendadas:
- Aderem, A., & Underhill, D. M. (1999). Mechanisms of phagocytosis in macrophages. Annual Review of Immunology, 17, 593–623.
- Desjardins, M. (2003). ER-mediated phagocytosis: A new membrane for new functions. Nature Reviews Immunology, 3(4), 280–291.
- Stuart, L. M., & Ezekowitz, R. A. (2008). Phagocytosis and comparative innate immunity: Learning on the fly. Nature Reviews Immunology, 8(2), 131–141.
- Swanson, J. A. (2008). Shaping cups into phagosomes and macropinosomes. Nature Reviews Molecular Cell Biology, 9(8), 639–649.
- Tollis, S., Dart, A. E., Tzircotis, G., & Endres, R. G. (2010). The zipper mechanism in phagocytosis: Energetic requirements and variability in phagocytic cup shape. BMC Systems Biology, 4, 149.
- Underhill, D. M., & Goodridge, H. S. (2012). Information processing during phagocytosis. Nature Reviews Immunology, 12(7), 492–502.
- Li, K., & Underhill, D. M. (2026). Phagocytosis: A process that shapes immune responses to engulfed meals. Nature Reviews Immunology, 26, 255–268.
Aprende administración paso a paso

ADMINISTRACION DESDE CERO