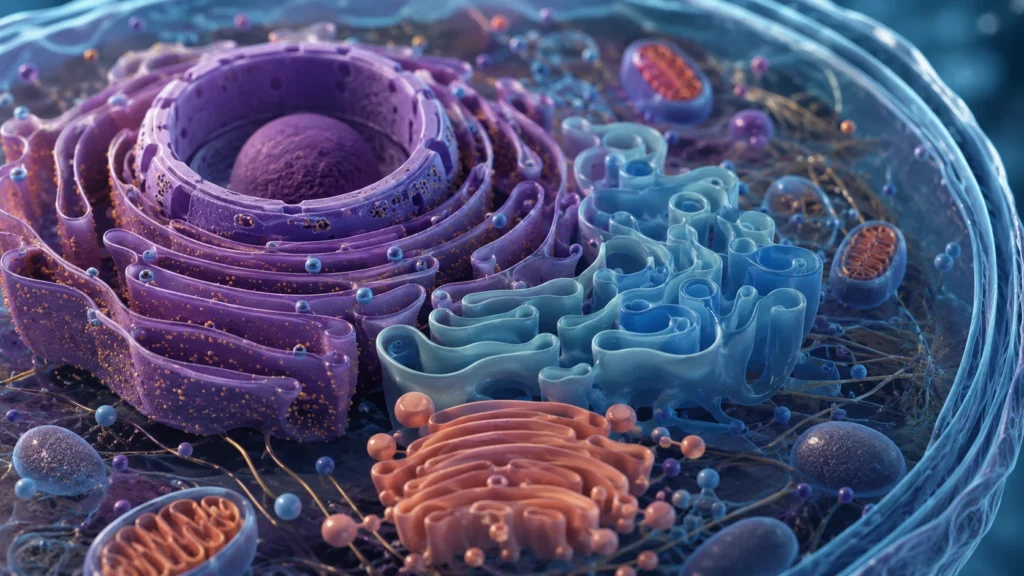
El retículo endoplásmico constituye uno de los sistemas membranosos más extensos y dinámicos de las células eucariotas. Se organiza como una red continua de cisternas aplanadas, túbulos ramificados y vesículas interconectadas que se distribuyen por casi todo el citoplasma y mantienen continuidad física con la envoltura nuclear. Esta continuidad estructural permite que el lumen del retículo endoplásmico funcione como un compartimento único especializado en síntesis, modificación, transporte y distribución de biomoléculas. La organización tridimensional del retículo endoplásmico no es aleatoria, sino que responde a requerimientos funcionales específicos de cada tipo celular. Las células con intensa síntesis proteica presentan grandes regiones de retículo endoplásmico rugoso, mientras que las células dedicadas al metabolismo lipídico o a la detoxificación poseen abundantes regiones de retículo endoplásmico liso.
La estructura del retículo endoplásmico está formada por membranas de bicapa lipídica semejantes a la membrana plasmática, aunque con una composición proteica especializada. Estas membranas delimitan un espacio interno denominado lumen o matriz endoplásmica, el cual posee una composición química distinta de la del citosol. El lumen contiene proteínas chaperonas, enzimas de plegamiento, sistemas de control de calidad proteica, proteínas fijadoras de calcio y múltiples complejos enzimáticos que participan en modificaciones postraduccionales. La separación física entre el lumen y el citosol permite crear microambientes bioquímicos especializados indispensables para procesos celulares altamente regulados.
La enorme superficie membranosa del retículo endoplásmico explica su papel central en el metabolismo celular. En hepatocitos y otras células metabólicamente activas, la superficie del retículo puede superar ampliamente la de la membrana plasmática. Esta expansión de membrana incrementa extraordinariamente la cantidad de enzimas y complejos moleculares que la célula puede alojar simultáneamente. Como consecuencia, el retículo endoplásmico se convierte en el principal centro intracelular para la síntesis de proteínas secretoras, síntesis de lípidos, ensamblaje de membranas, detoxificación de sustancias y regulación del calcio intracelular. Estudios morfométricos cuantitativos en hepatocitos demostraron que grandes proporciones del citoplasma están ocupadas por retículo endoplásmico rugoso y liso, reflejando la magnitud funcional de este orgánulo.
La conexión anatómica entre el retículo endoplásmico y la envoltura nuclear posee una enorme importancia funcional. La membrana externa nuclear es una prolongación directa de la membrana del retículo endoplásmico rugoso, de modo que ambos sistemas comparten el mismo espacio luminal. Esta continuidad facilita el transporte de proteínas recién sintetizadas, lípidos y componentes membranosos desde las proximidades del núcleo hacia regiones periféricas de la célula. Además, permite coordinar la expresión genética nuclear con los mecanismos de síntesis y procesamiento proteico localizados en el retículo endoplásmico.
La morfología del retículo endoplásmico depende de proteínas especializadas capaces de curvar, estabilizar y remodelar las membranas. Las regiones tubulares poseen elevada curvatura membranosa y suelen asociarse al retículo endoplásmico liso, mientras que las cisternas aplanadas se relacionan principalmente con el retículo endoplásmico rugoso. Esta diferencia arquitectónica no solamente representa una variación morfológica, sino también una adaptación funcional. Las cisternas amplias favorecen la unión de numerosos ribosomas y la síntesis masiva de proteínas, mientras que los túbulos facilitan reacciones metabólicas lipídicas y rápida distribución intracelular de membranas y moléculas.
El retículo endoplásmico rugoso recibe esta denominación debido a la presencia de ribosomas adheridos a la cara citosólica de sus membranas. Los ribosomas son complejos ribonucleoproteicos formados por ARN ribosomal y proteínas ribosómicas, cuya función consiste en traducir la información genética contenida en el ARN mensajero para sintetizar proteínas. La apariencia granular observada mediante microscopía electrónica se debe precisamente a la alta densidad de ribosomas asociados a las cisternas rugosas.
La unión de los ribosomas al retículo endoplásmico rugoso ocurre cuando una proteína en formación contiene una secuencia señal específica que dirige al ribosoma hacia receptores membranosos del retículo. Una vez acoplado, el ribosoma transloca la cadena polipeptídica naciente directamente al lumen reticular o a la membrana del retículo. Este mecanismo es fundamental porque garantiza que las proteínas destinadas a secreción, membranas celulares, lisosomas o compartimentos extracelulares entren inmediatamente en la vía secretora.
La síntesis proteica en el retículo endoplásmico rugoso implica procesos mucho más complejos que la simple formación de cadenas polipeptídicas. Dentro del lumen reticular ocurren plegamiento proteico, formación de puentes disulfuro, glucosilación inicial y ensamblaje de proteínas multiméricas. Las proteínas chaperonas luminales supervisan el correcto plegamiento molecular y detectan proteínas mal conformadas. Cuando una proteína no logra alcanzar su estructura funcional adecuada, es retenida y posteriormente degradada mediante mecanismos de control de calidad asociados al retículo endoplásmico. Este sistema es esencial para evitar acumulación de proteínas defectuosas potencialmente tóxicas.
Las células con intensa actividad secretora presentan retículo endoplásmico rugoso extremadamente desarrollado. Ejemplos clásicos incluyen células acinares pancreáticas, plasmocitos productores de anticuerpos y hepatocitos sintetizadores de proteínas plasmáticas. En estas células, las cisternas rugosas ocupan extensas regiones citoplasmáticas debido a la enorme demanda de síntesis y exportación proteica.
Por otra parte, el retículo endoplásmico liso carece de ribosomas adheridos y adopta predominantemente una organización tubular. Aunque comparte continuidad física con el retículo rugoso, sus funciones metabólicas son diferentes. La ausencia de ribosomas permite destinar mayor superficie membranosa a enzimas involucradas en metabolismo lipídico, detoxificación y almacenamiento de calcio.
Una de las funciones más importantes del retículo endoplásmico liso es la síntesis de lípidos. En sus membranas se localizan enzimas responsables de la biosíntesis de fosfolípidos, colesterol, triglicéridos y esteroides. Estos lípidos son indispensables para la formación de nuevas membranas celulares y para la producción de hormonas esteroideas. Las células endocrinas productoras de hormonas esteroideas, como las células de Leydig y las células corticales suprarrenales, poseen un retículo endoplásmico liso extremadamente abundante debido a la elevada demanda de síntesis lipídica.
El retículo endoplásmico liso también desempeña un papel esencial en la detoxificación celular. En hepatocitos, las membranas del retículo contienen sistemas enzimáticos como el citocromo P450, capaces de transformar compuestos liposolubles tóxicos en moléculas hidrosolubles más fáciles de eliminar. Medicamentos, alcohol, carcinógenos y metabolitos tóxicos son procesados mediante oxidación, reducción e hidroxilación en estas membranas. La exposición prolongada a ciertos fármacos induce expansión significativa del retículo endoplásmico liso hepático, aumentando la capacidad detoxificadora celular.
Otra función crucial del retículo endoplásmico es el almacenamiento y regulación del calcio intracelular. El lumen reticular actúa como reservorio de Ca2+, mantenido mediante bombas dependientes de ATP y proteínas fijadoras de calcio. La liberación controlada de Ca2+ desde el retículo hacia el citosol participa en señalización celular, secreción, metabolismo, apoptosis y contracción muscular. En células musculares, el retículo endoplásmico especializado denominado retículo sarcoplásmico regula directamente los ciclos de contracción y relajación mediante liberación y recaptación rápida de calcio.
La organización continua del retículo endoplásmico facilita el transporte intracelular de sustancias. Las proteínas y lípidos sintetizados pueden desplazarse a través del lumen o incorporarse a vesículas de transporte dirigidas hacia el aparato de Golgi. Esta capacidad de distribución convierte al retículo en la primera estación de la vía secretora celular. Sin este sistema membranoso integrado, la célula no podría coordinar adecuadamente síntesis, modificación y distribución de biomoléculas complejas.
Actualmente se reconoce que el retículo endoplásmico constituye además un orgánulo dinámico capaz de modificar continuamente su forma y distribución en respuesta a señales fisiológicas, estrés celular y demandas metabólicas. Las proporciones relativas de cisternas rugosas y túbulos lisos cambian según las necesidades funcionales de la célula. Esta plasticidad estructural permite adaptar rápidamente la capacidad biosintética y metabólica celular frente a variaciones ambientales o fisiológicas.

Fuente y lecturas recomendadas:
- Amar-Cortés, E. C., et al. (1989). Identification of the molecular components of rough and smooth endoplasmic reticulum membranes. Journal of Cell Biology, 108(2), 399–409.
- Baumann, O., & Walz, B. (2001). Endoplasmic reticulum of animal cells and its organization into structural and functional domains. International Review of Cytology, 205, 149–214.
- Loud, A. V. (1968). A quantitative stereological description of the ultrastructure of normal rat liver parenchymal cells. Journal of Cell Biology, 37(1), 27–46.
- Palade, G. E. (1975). Intracellular aspects of the process of protein synthesis. Science, 189(4200), 347–358.
- Schmucker, D. L. (1976). Age-related changes in hepatic fine structure: A quantitative analysis. Journal of Gerontology, 31(2), 135–143.
- Shibata, Y., Voeltz, G. K., & Rapoport, T. A. (2006). Rough sheets and smooth tubules. Cell, 126(3), 435–439.
- Voeltz, G. K., Rolls, M. M., & Rapoport, T. A. (2002). Structural organization of the endoplasmic reticulum. EMBO Reports, 3(10), 944–950.
- Westrate, L. M., Lee, J. E., Prinz, W. A., & Voeltz, G. K. (2015). Form follows function: The importance of endoplasmic reticulum shape. Annual Review of Biochemistry, 84, 791–811.
- Westrate, L. M., et al. (2015). The endoplasmic reticulum: Structure, function and response to cellular signaling. Cellular and Molecular Life Sciences, 72(1), 79–94.
Aprende administración paso a paso

ADMINISTRACION DESDE CERO